


研究内容
当教室では、各自のバックグラウンドをもとに、それぞれのメンバーが独自のテーマで研究を行っています。これらの研究のうち、代表的なものをご紹介します。
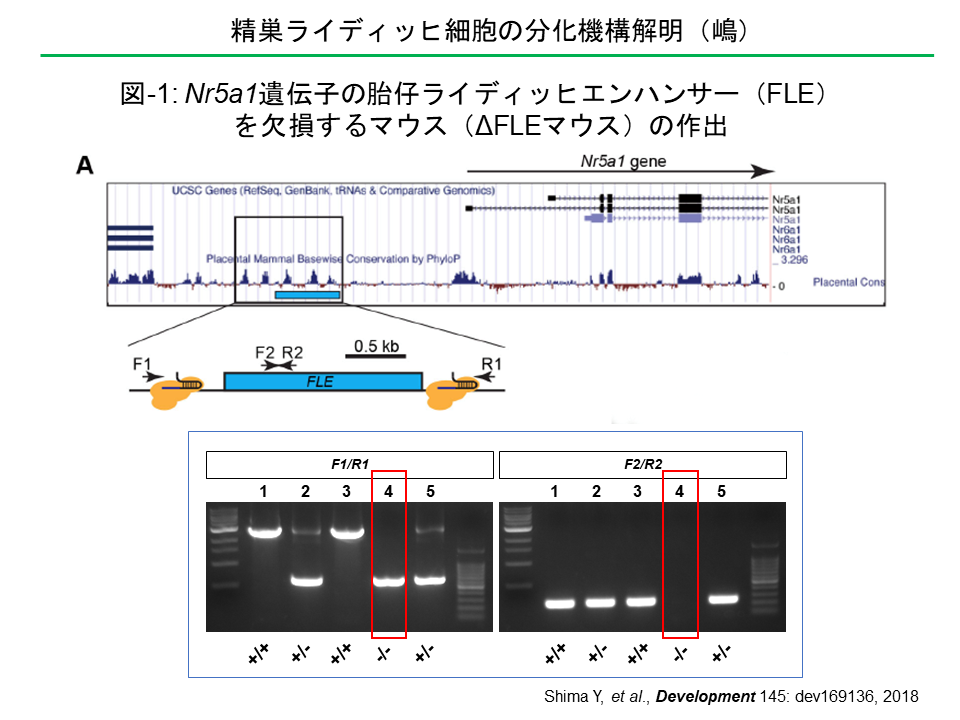
Nr5a1(Ad4BP/SF-1)遺伝子は、副腎、生殖腺、下垂体などの形成に必須の転写因子をコードしている。本遺伝子の発現は組織特異的エンハンサーによって制御されており、そのうち、胎仔の精巣に存在する胎仔ライディッヒ細胞における発現は、遺伝子上流のエンハンサー(Fetal Leydig Enhancer, FLE)により制御されている。FLEの機能を生体レベルで確認するために、ゲノム編集によりFLEを欠損するマウス(ΔFLEマウス)を作出した。
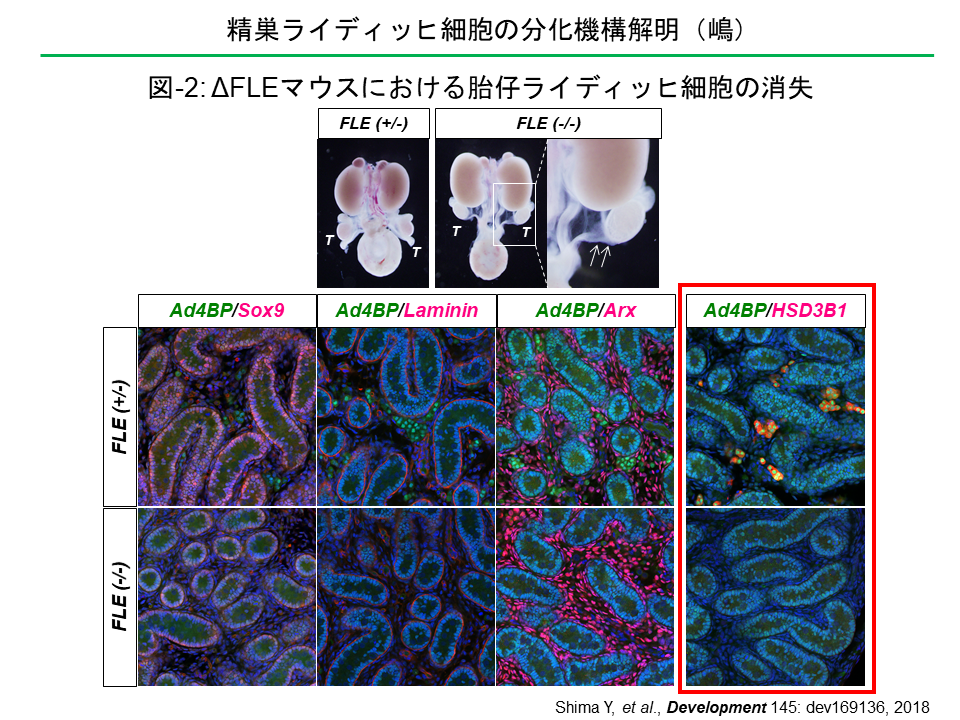
ΔFLEマウスの胎仔では、精巣が下降しておらず、輸精管が低形成であった(矢印)。免疫染色により精巣を解析した結果、Sox9陽性のセルトリ細胞、Lamininを含む基底膜、Arxを発現する間質未分化細胞は大きな影響を受けなかったのに対して、HSD3B1を発現する胎仔ライディッヒ細胞が完全に消失していた。この結果、胎仔精巣からのアンドロゲン産生が消失し、胎仔の雄化が阻害されていた。
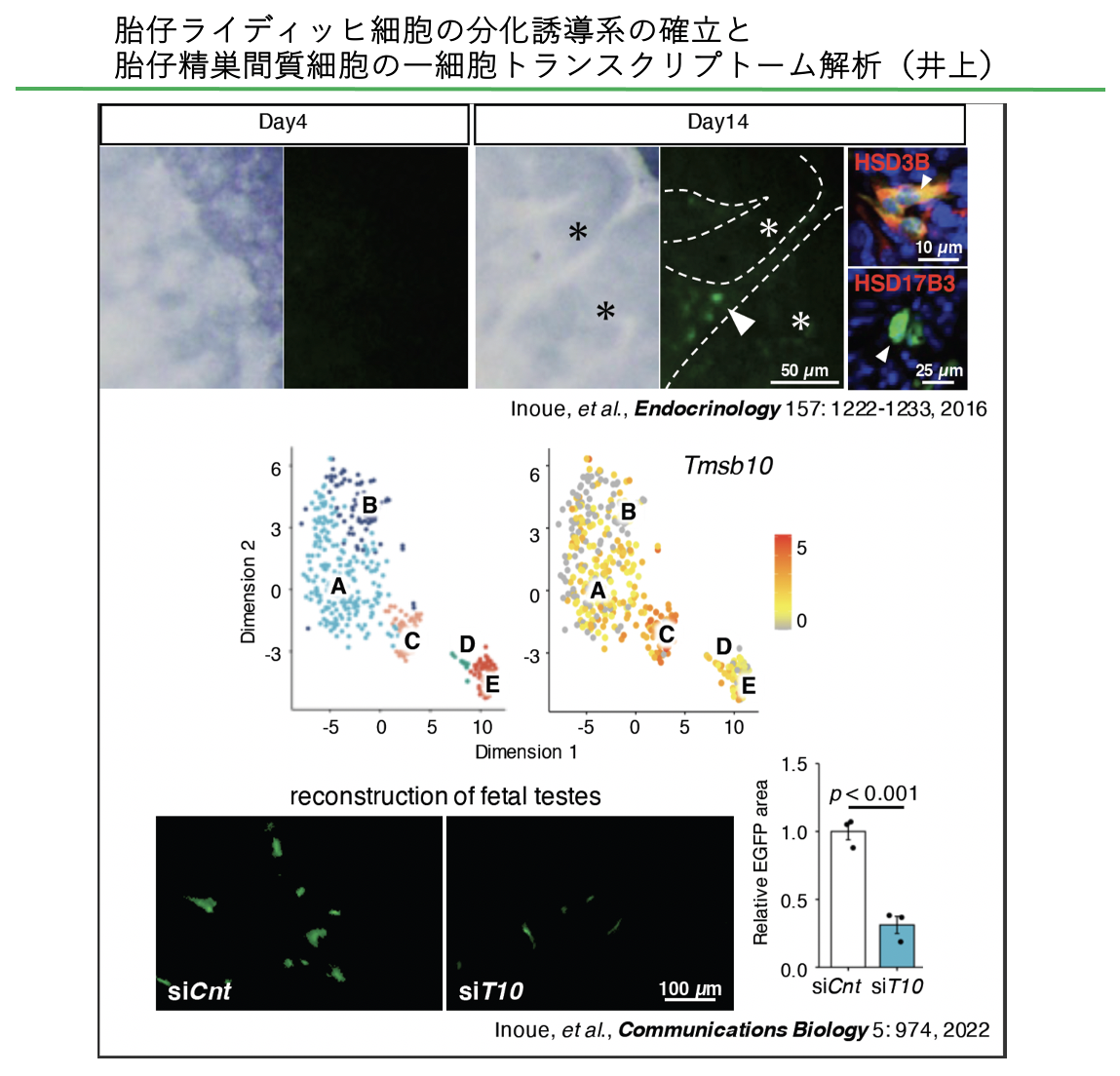
胎仔ライディッヒ細胞は細胞分裂をしないことが知られており、間質に存在する前駆細胞からの分化によってその細胞数が増加すると考えられている。しかし、胎仔ライディッヒ細胞の前駆細胞は同定されていない。我々は胎仔ライディッヒ細胞にEGFPを発現するマウスを作出し、胎仔ライディッヒ細胞に加え、EGFP弱陽性の間質細胞の単離に成功した。さらに、胎仔ライディッヒ細胞の分化を再現する再構築培養系を確立し、EGFP弱陽性の間質細胞に胎仔ライディッヒ細胞の前駆細胞が存在することを示した。さらに、胎仔ライディッヒ細胞と間質細胞の一細胞トランスクリプトーム解析から、前駆細胞と推測される細胞集団でTmsb10 遺伝子が高発現していることを見出した。その発現を抑制したところ、ヘッジホッグシグナル伝達が阻害され、胎仔ライディッヒ細胞の分化が抑制された。これらのことから、前駆細胞で一過性に発現するTmsb10が胎仔ライディッヒ細胞の分化の引き金となることが明らかになった。
